Do you know what people are never asking me?
"Hey Ben, how do I process proteomics data if I only have 3.5V of electricity and $25?"
My answer? You need a Raspberry Pi.
First of all -- I need to place credit where credit is due -- this is not an original idea. I first saw something like this at a place I visited that I'm pretty sure I can't say that I went to -- or what I saw there or what they were doing. So...I'm extremely unsure how to credit this, but this wasn't originally my idea -- thank you Dr. S.?
Second -- I need to credit the ladies in my life -- my 10 year-old niece for inspiring this project and Dr. Norris for getting it going "hey, lets all learn Python on Xmas break so Rhianna can rewrite and skip all the boring parts of her favorite video game!" (In action, above)
Third of all -- y'all do know about these little things, right?
This is a Raspberry Pi #3. Since my weird orangutan hands aren't good for scale, use the HDMI plug for your frame of reference. It is tiny (and there are smaller ones now! we're building some super cool little things around the house with ones that aren't much bigger than thumb drives!). It is essentially just a tiny PC motherboard with an HDMI output, a power supply, WiFi, Ethernet and 4 USB ports -- and that is about it. These things cost like $35, though one of ours we got for Black Friday all set up for $15 (though it didn't get a cool transparent case shown above ~$15)
Now -- I really had no idea what to expect coming out of this project. The Pi3 has a quadcore processor, but so do most modern cell phones!
What do you need?
A memory card (we got the maximum 32GB microSD that this board supports -- pushing this project to a whopping $65...which made me feel dumb, till I realized that's not unusually high for a video game these days!)
An operating system (I'm using Raspbian at first -- cause it looks JUST like Windows, LOL! Incidentally, the operating system comes with a bunch of video games --including Tetris! Win!)
Python (comes with Raspbian already installed -- plus things that teach you the language! But it is Python 2.0 and 3.0...complications remain to be seen!)
Okay -- so this thing runs Python and there are tons of Python tools that are out there in the world. Part of this exercise has been for Ben to finally (maybe) be able to evaluate some of them there Python tooly thingamajings. If I'm curious, I will honestly load up and check your R package (this is, however, very rare). If pressed because you did something super awesome -- yes, I will load up command line. But until now, I'll just have to take your word for it that you wrote some awesome Python code (or...honestly...not write about it...)
But there are awesome tools! Like:
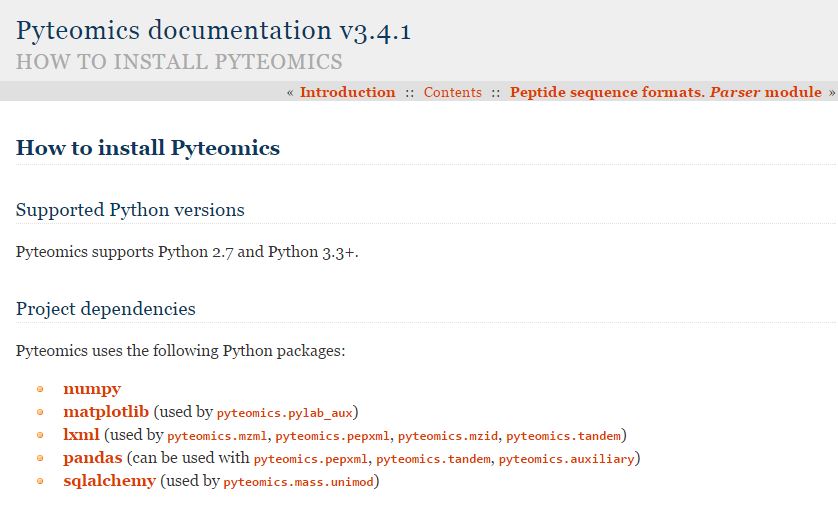
Pyteomics!
Looks great, and there are people doing stuff like this all the time. It remains to be seen whether I can run this on my handheld box. Maybe I'll get there.
But -- get this -- there are premade proteomics tools for Raspberry Pi already out there! The idea for this post isn't original in any way at all! Even better!
(These need to be screenshots from my Windows PC, I'm not energetic enough to do this from the Pi and email them...)
There are HUNDREDS (maybe thousands!) of awesome scientific tools already developed to run on these little boxes.
Check this list. I know I was amazed. Everything from DNA sequence alignment to imaging software to -- you guessed it, mass spectrometry data processing!
The first thing that pops up for ctrl+F "proteomics" is
Mmass! It appears to have all the same functions on this little box as it does on Windows.
But -- check this out -- those forward thinkers at the
GPM made an X!Tandem Raspberry Pi package!
Okay -- so I found a search engine (even better, one that I'm familiar with!) and -- well, this is a long term rainy day project (that has been somewhat hampered by the fact the Pi has to be really close to it's WiFi source for me to download stuff) -- so more on this topic later -- eventually...probably...my motivation isn't quite where it was when I started writing this, but I still feel like hitting the "Publish" button. I had no idea this was such a legit scientific tool that would be so easily accessible for a non-programmer!